

2016. február 28. - A fák vízszállításáról újszerűen
A ForestPress hírportál hasábjain 2015. július 28-án cikket írtam „A pulzáló rendszer – a fák vízszállításáról újszerűen” címmel. Emlékeztetőül röviden ismertetném a pulzációs elmélet lényegét.
Párolgáskor a lombkorona irányába nem szívás, hanem nyomás történik a párolgás miatti hőelvonás következtében előállt vízcső keresztmetszet csökkenése miatt. Éjszaka a párolgás szünetelésekor beindul egy hő-kiegyenlítődési folyamat, ami visszaállítja a cső eredeti keresztmetszetét. Ez szívást generál a gyökerek irányába, és felszívja a vizet a talajból. Ennél fogva a cső teljes keresztmetszete pulzál, ami lehetővé teszi az állandó erőhatás miatti oldatszállítást a csőben. A cikkel kapcsolatban felmerült egy érdekes kérdés. Mivel magyarázható, hogy a téli lombtalan állapot után tavasszal be tud indulni az élet. Ne felejtsük el, hogy ekkor még sem párolgás, sem fotoszintézis sincs. A fák „vegyi üzemei” a rügyfakadási időszakra előállítják azokat a vegyületeket, amiből a még rügyben rejtőző hajtásrendszer felépíthető.
A kérdés a következő: Vajon hol rejtőznek az építőanyag többletvegyületei, és milyen erő tudja ezt a rendeltetési helyére juttatni? Megpróbálom ezt a pulzációs elmélettel megmagyarázni.
Véleményem szerint ősszel a színesedő levelek úgy esnek le a fáról, hogy a nyomás benne marad a rugalmas szállítórendszerben. Szeretném ezt egy gyakorlati példával megvilágítani, majd a fák esetében ezt bizonyítani. Bizonyára ismert kedves olvasóim előtt az a jelenség, amelyet kiskertünkben slaggal (rugalmas gumicső) történő locsolásánál tapasztalunk. A cső rugalmas, benne nyomás van. A locsolás befejeztével a locsolópisztoly billentyűjének lezárásával a nyomás bent marad a csőben többletvízként. Erről könnyen meggyőződhetünk úgy, hogy a cső lezárása után leállítjuk a szivattyút, majd ezek után újra szabaddá tesszük a vízcső végét a pisztoly billentyűjének segítségével. Tapasztaljuk, hogy a többletvíz, nyomással „kilövell” a cső végén.
Ugyanez a jelenség történik a fánál is. A fának nem is áll módjában másként lezárni a csövek végét, csak nyomás alatt, hisz a „motor” a pulzáló cső, ha egyre csökkenő mértékben is, de felfelé nyomva működik. Bizonyíték: Tél végén, amikor ez a nyomás a csőből részben kiszabadul, hisz a vegyületekkel a kapcsolatot meg kell teremtenie (bár ez a kapcsolat egész évben megvan), akkor azt tapasztaljuk, akár a legmagasabban lévő ágak lemetszésénél is, hogy a nyomás alatt lévő többlet cukros vegyület kifolyik a fából. Amíg nincs transzspiráció és fotoszintézis, ez a jelenség biztosítja a rügyekben szunnyadó hajtásrendszer kialakulását.
Régebben ezt a gyökérnyomásnak nevezték. Mára már a tudomány bebizonyította, hogy gyökérnyomásról nem lehet szó, mert az egy nagyon kicsi érték. A csíranövényekben a lomblevelek kifejlődéséig lehet szerepe. Működés-technikailag azért sem beszélhetünk itt gyökérnyomásról, mert egy mechanizmus, ha működik, akkor a víznek állandóan folynia kellene. A fent leírtak a pulzációs elmélet egyik bizonyítékát jelentik.
Nappali szobánk elé délnyugati kitettségbe egy fügefát ültettem, melyet Majer Antal (soproni egyetemi – szerk.) professzor hozott nekem ajándékba arácsi birtokáról. Ez a fügefa egy évben kétszer terem. Nyár derekán és októberben. Az októberi érésű fügének van egy jellegzetes tulajdonsága, mégpedig az, hogy még hosszú, meleg ősz esetén is a termésének kis hányada érik be. Ami beérik, az viszont szinte egyik napról a másikra hirtelen megnövekszik. A feltételezésem az volt, hogy a fügeérésnek ebben a stádiumban nagy a növény vízfelhasználása a hirtelen termésérlelés miatt. Ekkor letörtem a fügefáról egy levelet közvetlen a gyorsan duzzadó füge termése mellől, a levélnyéllel együtt, majd a levélnyelet az ég felé fordítottam. A levélnyél alapján oldat szivárgott ki, majd csepp formájában végigfutott a levélnyélen. Nem kell sokat bizonygatnom, hogy ilyen jelenség kizárólag felfelé történő nyomás esetén valósulhat meg. Ez a nyomóerő feszíti ki a leveleket, amelyek ettől a feszes állapottól oly szépek és szemet gyönyörködtetőek. Ha csak szívásról beszélünk, akkor ez a feszes állapot nem jöhetne létre. Ugyanezt a kísérletet éjszaka is elvégeztem. (A pulzációs elmélet szerint a zárt sztómáknál a vízcső visszaállítja eredeti keresztmetszetét, ami szívást generál a gyökerek irányából.) A levélalapon ekkor csorgást már nem tapasztaltam. Az elmélet egy másik bizonyítékát Béres Csilla kísérlete szolgáltatta, aki éjszaka zárt sztómáknál is tapasztalt izotópos kísérlete során egy lassú felfelé történő áramlást. Meg is jegyezte: „a párolgás felfelé történő húzóerejével ez a jelenség nem magyarázható”. A pulzációs elmélettel viszont igen. Hiszen éjszaka, amikor a gyökerek irányából szív a növény, akkor is kell lennie egy lassú felfelé történő áramlásnak duzzadó csövekkel.
Gyakran lehet olvasni növényélettani könyvekben, hogy a gyökérnyomásnak van egy napi ciklusa és ez reggel a legmagasabb érték, mielőtt a gázcsere nyílások kinyílnak, és a párologtatás megkezdődik. Vajon miért? A fiatal vízcső a két, már megszilárdult merev hengerpalást között helyezkedik el. Az éjszakai gyökér felőli szívás esetén a cső egész éjjel duzzad. A legnagyobb duzzadtságot sztómanyitáskor éri el, amikor a cső elkezd felfelé nyomni. A cső nyomását véleményem szerint nem tudjuk mérni, de az ugyanakkora nagyságú és ellentétes irányú ellenerőt, amit a két merev hengerpalást fejt ki az embrionális állapotú különböző feladatok ellátására hivatott sejtekre, azt igen.
Tekinthetjük ezt is egy közvetett bizonyítéknak. Tehát itt sem gyökérnyomásról van szó (mint már korábban írtam, kiderült róla, hogy ez egy nagyon kicsi nyomás).
A pulzálás féregmozgásai
Felmerül a kérdés: valóban elegendő erő rejlik a pulzálásban ahhoz, hogy akár egy nyolc méter magas fa csúcsába vizet juttasson?
A probléma az, hogy a vízcsőben egy nyomásgrádiens alakul ki a fa csúcsától a gyökerek végéig. Értelemszerűen ez a hidrosztatikus nyomás a fa csúcsánál a legkisebb és a fa legalacsonyabban fekvő gyökéri résznél a legnagyobb érték.
A hidrosztatikus nyomás: a folyadék sűrűségének, a gravitációs gyorsulásnak és a folyadékoszlop magasságának a szorzata.
A fent említett nyolc méteres magasság esetében a földfelszíni vágáslapon ez a hidrosztatikus nyomás 80 KPa (kilopaszkál). Mi tehát a teendő, hogy ettől a magasságfüggvényű változó nagyságú hidrosztatikus nyomásból fakadó problémától megszabaduljon a növény? A nehézséget az okozza, hogy változó nagyságú nyomó, illetve szívóerőre van szükség és ennek legnagyobb szélső értékével még kis magasság esetében sem tud megbirkózni a fa.
Nézzük meg, hogy az épületgépészek hogyan juttatják fel akár háromszáz méternél is magasabb felhőkarcoló felső emeletére a vizet. Elvileg két megoldás létezik.
Létrehozunk egy egyenszilárdságú csövet egy nagyon erős vízpumpával és a szerelvényeket nyomáscsökkentőkkel védjük. Ilyen konstrukció tudomásom szerint nem készült.
Az alkalmazott megoldás a vízvezetékrendszer leszakaszolása puffer tárolók segítségével. A növény vízcsöve is ezt a módszert választotta. A leszakaszolás darabszámának arányában csökken a hidrosztatikus nyomás, de elérhető az is, hogy a szükséges oldalirányú erő oly mértékűre változzon, ami lehetőséget teremt ahhoz, hogy általa a leszakaszolt csőben felfelé nyomás történhessen.
A másik érdekes és fontos kérdés, hogy vajon ebben az esetben milyen pulzálási mozgásforma alakul ki. Vizsgáljuk meg, hol találunk leszakaszolásokat és ott milyen a mozgásforma. Az ember vékonybelében ilyen a mozgás, de a gyűrűsférgek is hasonlóan mozognak, sőt a pocok is így mozog, mikor állkapcsát kiakasztva lenyeli a kígyó és továbbmozgatja bélrendszerében. Talán már sejtjük. Egy perisztaltikus mozgásformával (féregmozgás) állunk szemben. A fák vízcsövének esetében ez egyedi, sok tekintetben eltér a többi perisztaltikus mozgástól, de sok a hasonlóság is. A vékonybél perisztaltikus mozgásától annyiban tér el, hogy ott van egy szegmentációs (leszakaszoló) mozgás is, mely az idő függvényében előrehalad, és a megemésztett anyag forgatásában, keverésében is szerepet játszik. A fák vízcsövénél szegmentációs mozgásra nincs szükség, mert azok a szelepek által eleve leszakaszoltak. Vizsgáljuk meg ezeknek a leszakaszolt szegmentumoknak a mozgását, tárjuk fel a szelepek zárásának, nyitásának a mechanizmusát.
Fny = nyomóerő, Fe = ellenerő (több erőből áll, ezért nem ábrázoltam)
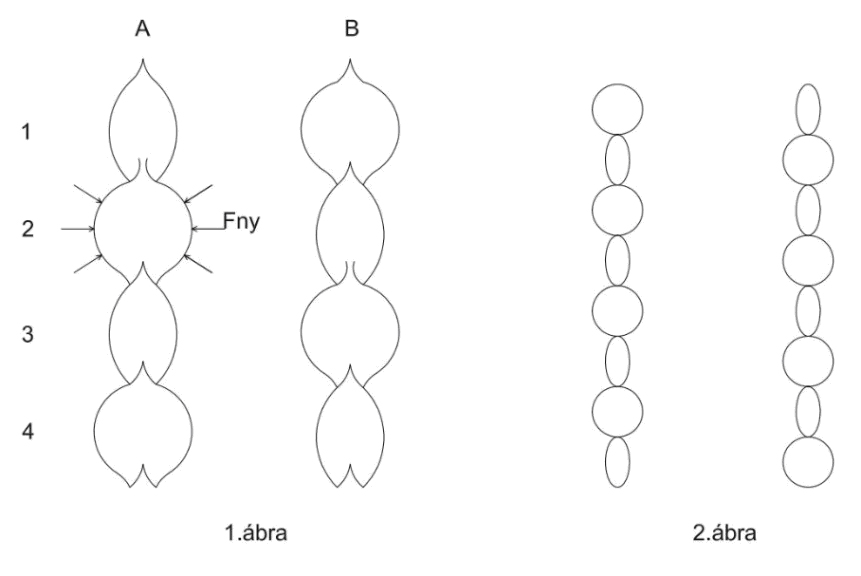
A szelepek zárásának, nyitásának a mechanizmusa
1A ábra: 2. számú szegmentumból vizet nyom a növény a felette lévő 1. számú szegmentumba. A szelepnyitás feltétele: Fny > Fe. A nyomási folyamat közben az ellenerőnek meg kell nőnie ahhoz, hogy a szelep bezáródjon. Abban a pillanatban, amikor Fny < Fe, záródhat a szelep. Ezután megkezdődhet a nyomás a 3. szegmentumból a felette lévő előző nyomással lecsökkentett vízmennyiségű 2. számú szegmentumba. (1B ábra szemlélteti az eredményeket). A szelep nyitás-zárás előfeltétele egyezik az előbbivel. Ez a folyamat folyton-folyvást ismétlődik és megkapjuk az 1. ábrán szemléltetett perisztaltikus mozgásformát. Abban az esetben, ha a 2. ábrát egymás után ráragasztanánk egy hengerpalástra úgy, hogy az ábrák záródjanak a hengerpaláston (az első érintkezzen az utolsóval), majd a hengerpalástot függőleges tengelye körül megpörgetnénk, megkaphatnánk a perisztaltikus mozgás mozgó változatát. Animáció szempontjából nem lehetséges, hogy a 2. ábra egy fraktál elem, melynek forgatással történő ismétléseivel a mozgásforma megjeleníthető? Ezzel a perisztaltikusan pulzáló mozgásformával már lehetősége van a csőben áramló víznek a legmagasabb fák csúcsába is feljutni. Az éjszakai szívás esetén a perisztaltikus pulzálás megmarad, mivel a cső duzzad, a mozgás lassabb és egyenletesebb, de ekkor is felfelé áramlik.
Keressünk bizonyítékokat a féregmozgásos pulzálásra, nehogy a fent leírtak egy tőmelletti öreg erdész hagymázas fantáziájának tűnjenek! Béres Csilla izotópos kísérletére szeretnék utalni, amikor az időfüggvényű sebességmintázatokat elemzi. Ő a felfelé történő áramlásnál rövid ideig tartó kiugró csúcsokról beszél, melynek eredője egy gyors nappali áramlás lehet. Béres Csilla cikkét folytatásos formában az Élet és Tudomány 1999/50. és 1999/51. számában ismertette „A fák vízforgalma” címmel. Sajnos ezekben a cikkekben időfüggvényű sebesség grafikonokat nem találtam, csak nagyon részletes mozgáselemzések leírását.
Béres Csilla időközben Debrecenből áttette székhelyét Szombathelyre, ahol immáron hordozható CT (kompútertomográf) segítségével folytatta a fák vízszállítási kutatásait. Az elért eredményekről „A fafajok vízforgalmának összehasonlítása” címmel a Természet- Műszaki és Gazdaságtudományok alkalmazása 7. nemzetközi konferenciáján (Szombathely, 2008. május 17.) számolt be. A kutatócsoport tagjai: Béres Csilla, Németh László, Zentai Zoltán NYME TTMFK Kémiai és Környezettudományi Tanszék kutatói voltak.
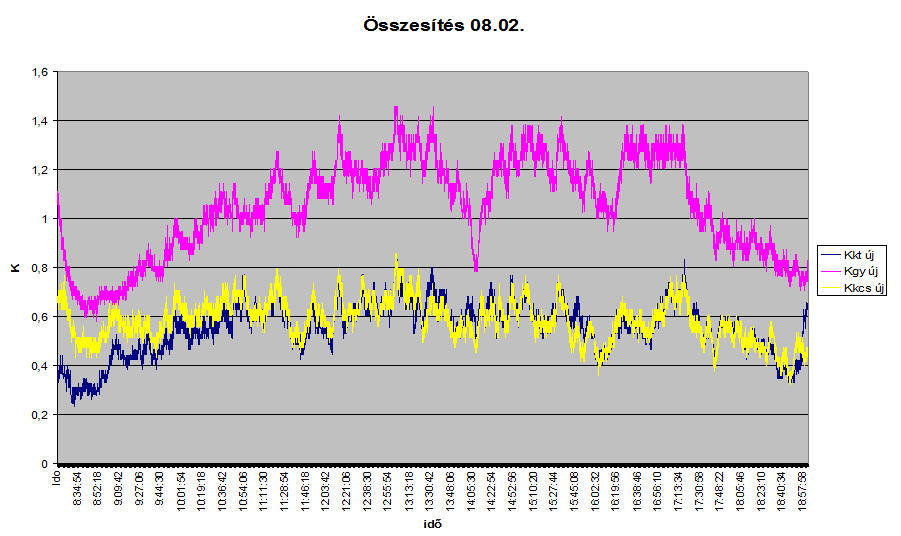
3. ábra: Csertölgy, kocsánytalan tölgy és gyertyán törzsében mért vízáramlás locsolás utáni napon.
A függőleges tengelyen K=0206U a 0,814-ezrediken, ahol U az áramlási sebesség m/perc. A 3. ábrán a csertölgy, kocsánytalan tölgy és a gyertyán törzsében mért vízáramlás időfüggvényű sebességmintázatai látszanak egy locsolás utáni napon. Ugyan az látszik, amit az izotópos kísérletnél is leírt. Az áramlás nem egyenletes, hirtelen gyors és lassú áramlások követik egymást. Vajon miért? A féregmozgásos pulzációs elmélet szerint a víz egyik szegmentumból áramlik a felette lévőbe. Kialakulhat-e itt egyenletes mozgás? Véleményem szerint nem. Amikor a szegmentum oldalirányú ereje felnyitja a felette lévő szelepet, akkor a víz meglódul, majd lelassul. Tulajdonképpen arról van szó, hogy a felfelé haladó víz kinetikus energiájának egy része elraktározódott a rugalmas szelepbe, melyre hidrosztatikus, és a rugalmas rendszerek nyomása is hat, majd a szelep felnyílásakor ennek a kinetikus energiának egy része felszabadul egy hirtelen gyors, majd folyamatosan lassuló vízmozgásformát előidézve. Ez hasonlítható az íj felajzásához. Kinetikus energiát fektetünk be, melyet a rugalmas íj felajzás után egy ideig tárol, és amikor ezt felszabadítjuk (kilőjük a nyílvesszőt), ennek a betárolt kinetikus energiának egy részét visszanyerjük kinetikus energiaként. Morfológiai bizonyíték: Minden vízcsőről készített ábrán látható a leszakaszoltság, de ez nem is alakulhatott volna másként, hisz különböző hosszúságú vízszállító sejtek rendeződtek egy függőlegesen kapcsolódó alakzatba. A CT által készített időfüggvényű sebességmintázat tehát visszatükrözi a szegmentumról szegmentumra áramló víz mozgásformáját.
Egy kis bionika
Nem tehetem meg, hogy néhány gondolat erejéig ne érintsem a bionikát, ezt a modern tudományt, hisz írásomban sokszor oda-vissza élő és élettelen rendszereket hasonlítottam össze. A bionika olyan új, több szaktudományt átfogó tudományág, amelynek célja az élő természetben kifejlődött megoldások átültetése a műszaki gyakorlatba, abból a megfontolásból is kiindulva, hogy a természetben fennálló természetes kiválasztódás az optimális megoldásokat jelenti egy-egy problémára. A természet ingyen mindenféle ellenszolgáltatás nélkül bocsátja az ember rendelkezésére találmányait. Nyugodtan kijelenthetjük, hogy a bionika is a természet adományainak haszonélvezője.
A növényvilág találmányairól Móra Ferenc így írt 1933-ban egy könyvismertetésben: „A növények a legnagyszerűbb kémiai gyárosok – panamák nélkül – nekik van a legtökéletesebb vízvezetékrendszerük, ők a legügyesebb szerelők és legzseniálisabb építészek, s már akkor traverzekkel dolgoztak és pillérekkel, amikor az embermag még el sem volt vetve a tengerek iszapjában.”
„Nincs az emberi technikának, mechanikának olyan találmánya, ami ne lenne jelen a növények világába.” – mondta Greguss Pál.
A bionika fő alapelve: Hasonló rendszerek hasonlóképpen működnek. Sokat elmélkedtem azon, vajon miből fakadhat ez a hasonlóság működéstechnikai szempontból. Talán a megoldás kulcsa a működéstechnikát előidéző mechanizmusokban rejlik? A mechanizmus lényegét a következőképpen lehetne megfogalmazni. A mechanizmus olyan önmagába záródó szimmetrikus mozgásforma, melyet az ellentétesen működő hatások tartanak szimmetrikus állapotban.
Térjünk vissza a fák vízszállító csövéhez, ehhez az élő sejtek függőleges összekapcsolódása által szerveződött csodálatos organikus képződményhez. Fellelhető ellentétes hatások: szívás - nyomás, keresztmetszet-növekedés – keresztmetszet-csökkenés, felmelegedés – lehűlés, szelepnyitás – szelepzárás.
Ha a felsorolt ellentétpárokból egy is kimarad, vagy az ellentétpárok ellentéteiből egyik is hiányzik, akkor az organikus rendszer mechanizmusa működésképtelenné válik.
A vizet szállító vízszivattyúknál ilyen sok ellentétpárról nem beszélhetünk. A vízszivattyút nem készítette fel az ember arra, amire a növény vízszállító rendszerének fel kellett készülnie. Alkalmazkodni a változó klímához, a változó diszponíbilis (rendelkezésre álló – szerk.) vízkészletekhez. Ezért az organikus rendszerek mechanizmusai sokkal kifinomultabbak.
Rengeteg vízszállító rendszert ismerünk, amelyek különböző mechanizmusok segítségével szállítják a vizet.
Még perisztaltikus szivattyú is létezik, mely szinte leutánozza a vízszállító cső perisztaltikus pulzálását.
Közös jellemzőjük: minden szivattyúnál van szívás és nyomás. Van még egy közös, valamennyi szivattyúra jellemző tulajdonság. A légtérbe a vizet minden esetben a nyomási szakasszal juttatják ki. Miért pont a fáknál lenne ez másképp? Török András