

2017. április 9. - A ForestPress elektronikus hírportálon 2015. július 28-án A pulzáló rendszer – a fák vízszállításáról újszerűen, majd 2016. február 29-én A pulzáló rendszer féregmozgásai – a fák vízszállításáról újszerűen címmel jelentek meg cikkeim.
Olvasóim részéről igényként merült fel írásaim kiegészítése az alábbiakkal:
A jelenleg elterjedt elmélet ismertetése és ennek egyértelmű cáfolata
Azon pontok felkutatása, ahol az élettelen és az élő rendszer működése kapcsolódik egymáshoz, amelyek mögött nem csak fizikai, mechanikai, hanem élettani, biokémiai és biofizikai folyamatok húzódhatnak meg.
A két korábbi cikkem összeszerkesztésével, az említett hiányosságok pótlásával, valamint az azóta napvilágot látott rendkívül érdekes, a fák alvómozgásának magyarázatával kiegészítve adom közre írásomat. Íme…
Növényanatómiából elég jól ismerjük, hogy a kambium osztódó szövetei hozzák létre a fatest belseje felé a különböző feladatok elvégzésére hivatott fasejteket, így a vízszállításra specializálódott sejteket is, melyeknek egy része összekapcsolódásuk által vízszállító csövekké alakul át.
A következőkben kizárólag a hosszú távú szállítással kívánok foglalkozni, amikor a víz a csövekben, az úgynevezett xilém elemekben mozog. (Az edényes növényekben a xilém vagy farész a két szállítószövet egyike, a floém vagy háncsrész a másik.)
A jelenleg elterjedt elmélet és ennek cáfolata
Az ismert vízszállító elméletek csak kis eltéréssel különböznek egymástól, lényegi eltérések nem tapasztalhatók.
Ennek rövid leírása a következő:
A xilém nedv mozgásának legfontosabb hajtóereje a víz párolgása a mezofil (levélalapi) sejtek felületéről a légkörbe. A párologtatás hatására a mezofil sejtek falán milliónyi apró meniszkusz (a kapilláris folyadékoszlop görbült felszíne) jelenik meg. Ezek felületi feszültsége negatív nyomást, illetve húzóerőt gyakorol a xilémre, ami felhúzza a vizet a gyökerekből és a talajból. A vízszállítás alsó mozgatójaként a gyökérnyomást szokták megemlíteni. Ha a gyökérsejtek vízpotenciálja negatívabb a talajénál (pontosabban a talajoldaténál), általában az oldott anyag magas koncentrációja miatt a víz ozmózissal beléphet a talajból a gyökérbe. Ez pozitív nyomást hoz létre, ami a xilémnedvet a levelek felé mozgatja. (Megjegyzés: egyes szerzők a szakirodalomban a gyökényomásnak alárendelt szerepet szánnak.)
Felmerül a kérdés: működhet-e a fában a vízszállítás az ismertetett módon?
Egy rendszer akkor működik a célnak megfelelően, ha működőképes a mechanizmusa, létezik ehhez illeszkedő megfelelő nagyságú és állandóan ható erő úgy, hogy, szerkezeti elemei ezt képesek elviselni.
Vizsgáljuk meg ezeket külön-külön.
- Az egyik említett erő a hajszálcsövesség, idegen szóval kapilláris hatás vagy kapilláris effektus, a folyadék azon tulajdonsága, hogy képes szűk keskeny térben a gravitációs erő ellenében is mozogni. Ha a cső átmérője elég kicsi, akkor a felületi feszültség, valamint a cső közötti adhéziós erők együttes hatása felemeli a folyadékot a kapillárisban. A kapilláris emelés azonban a természetben csupán egy méter körüli érték, ennél nagyobb magasságba a kapillárisokban a víz nem tud feljutni. Az ozmózisból származó gyökérnyomás (nyomóerő) lehet kicsi és lehet nagy is, teljesen mindegy, folyamatos vízszállításra alkalmatlan, mert az erő csak az általa működtetett mechanizmusa révén képes folyamatos munkát végezni, például vizet szállítani (később erre részletesen kitérek).
- A fák vízszállítási mechanizmusa a folyadékot szállító vízszivattyúkéhoz hasonlítható. Ezeknek a szivattyúknak a vízszállítás módját tekintve két típusuk ismert. Az azonos időfázisban szívó-nyomó vízszivattyúk, ilyen például a közönséges szivattyú, ahol az erőforrás által működtetett vízkerék két részre osztja a rendszert. A vízbe érő szívócsonk és a vízkerék egyik fele között a szívott vízszakasz. A vízkerék túloldala és a szabad légtér között pedig a nyomott víz található.
A szivattyúk másik típusa, amikor a szívás és a nyomás nem azonos időben történik, hanem késleltetve. Ezek az úgynevezett eltolt időfázisú szivattyúk (például az orvosi fecskendő). Először felszívjuk a folyadékot, majd a következő lépésben kinyomjuk.
Mindkét szivattyútípusra igaz: Először szívunk, majd a felszívott folyadékot kinyomjuk. A szívott és a nyomott vízszakasz határozottan elkülönül.
A leírtakból következik: vizet szállítani (ez organikus rendszerekre is igaz) csak szívással vagy csak nyomással nem lehet. Nem lehet vizet szállítani úgy sem, hogy alulról nyomok, felülről szívok (ebben az esetben a szívott és a nyomott vízszakasz nem tud elkülönülni). Mivel nincs a rendszerben állandó erőhatás által működtetett mechanizmus, így a csövekben víz nem áramolhat (a későbbiekben ezt is részletesen elemzem).
- Párolgásnál a csőnek víztelítettnek kell lenni, mert párolgás csak ebben az esetben valósulhat meg. Erőegyensúlyi feltétel bármely magasság esetében: a lefelé irányuló gravitációs erőnek egyensúlyban kell lenni a vele ellentétes irányú szívóerővel. Ahhoz, hogy egy bizonyos magasság esetén víz jusson a levelekbe, az erőegyensúlyt létrehozó szívóerőnél egy kicsivel nagyobb erő szükséges. Ha ezt egy gyakorlati példával akarom megvilágítani, például egy középmagasnak tekinthető 20 méteres bükkel, akkor ez azt jelenti, hogy a 20 méteren lévő levélben két atmoszféránál egy kicsivel nagyobb szívóhatásnak kell lenni. Két atmoszféra nyomással egy középkategóriás autó kerekét keményre lehet pumpálni.
A levél finom szerkezeti elemei ennek a fordítottját, vagyis a mínusz két atmoszféra szívást hogyan képesek elviselni? Ez teljes képtelenség. Nem beszélve arról, hogy 100 m magas fa esetén ennek ötszörösét, mínusz 10 atmoszférát (-1 Mpa-t) kellenne kibírniuk ugyanazon felépítésű levelek belső sejtjeinek.
Az elvégzett elemzésből kitűnik, hogy az eddig megismert vízszállítási elmélet hibás. Hibás azért, mert nincs benne a mechanizmushoz illeszkedő kellő nagyságú erő. Hibás a mechanizmusa, mert folyadékot úgy nem lehet szállítani. A legnagyobb ellentmondást az jelenti, hogy nagy magasság esetén a levél szerkezeti elemei szilárdságtani szempontból a keletkezett szívóhatást képtelenek elviselni.
Ha nem így, akkor vajon hogyan szállítják a vizet a fák?
A pulzáló rendszer működési elve
A levél felszínén igen kisméretű nyílások (sztómák) találhatók. Ezek a sztómák, melyek többnyire a levél fonáki oldalán helyezkednek el, gondoskodnak a légcseréről és a párologtatásról. Párologtatáskor a sztómák nyitva vannak. Magasabb energiaszinten vízmolekulák távoznak a légtérbe. Következésképpen a csőben maradó víz lehűl (hőelvonás jelensége). Mivel organikus rendszerről van szó, a vízcső a lehűlésre keresztmetszet- csökkenéssel válaszol. Ebben az esetben pedig nem szívásról, hanem a légtér felé irányuló nyomásról beszélünk. A cső csak egy bizonyos határig tud szűkülni. Az összehúzódás befejeztével bezárul a légzőnyílás. Megszűnik a párolgás és a hőelvonás. Beindul egy hőkiegyenlítődési folyamat, ami szép lassan visszaállítja a cső eredeti keresztmetszetét. Ez a keresztmetszet-növekedés pedig szívóerőt generál a gyökerek irányába, ami felszívja a vizet a talajból. Hasonlóan az orvosi szemcseppentőhöz, amikor is a benyomott, leszűkített keresztmetszetű, henger alakú, rugalmas gumit elengedve folyadékot szívunk fel a csőbe.
A víznyomáshoz (sztómanyitás – párolgás) szükséges nyomóerőt a párolgás miatti hőelvonás következtében létrejött, az egész csőhosszra kiterjedő keresztmetszet-csökkenés állította elő. A gyökerek felé irányuló szívóerőt, ami az eredeti keresztmetszetet hivatott helyreállítani, a hőkiegyenlítődés során keletkezett erő biztosítja. Jelen esetben a szívás és nyomás fázisban a cső teljes felülete pulzál, hisz magas fáknál nagy erőkifejtés szükséges.
Az elmondottakból érzékelhető, hogy a vízszállításnak ez az újfajta mechanizmusa csak fiatal, elsőéves, még összehúzódásra képes csövek esetén működhet. A növényanatómiával és élettannal foglalkozók megfigyelték, hogy a második évben a csövek már rugalmatlanok, bennük vízszállítás nem, csak oldatraktározás folyik. A többéves csövekben azért nem folyhat vízszállítás, mert a vízcső vége nincs közvetlen kapcsolatban a levéllel.
A pulzálás féregmozgásai
Felmerül a kérdés: valóban elegendő erő rejlik a pulzálásban ahhoz, hogy akár egy nyolc méter magas fa csúcsába vizet juttasson?
A probléma az, hogy a vízcsőben nyomásgradiens alakul ki a fa csúcsától a gyökerek végéig. Értelemszerűen ez a hidrosztatikus nyomás a fa csúcsánál a legkisebb és a fa legalacsonyabban fekvő gyökéri részénél a legnagyobb érték. (A hidrosztatikus nyomás a folyadék sűrűségének, a gravitációs gyorsulásnak és a folyadékoszlop magasságának a szorzata.)
Nyolcméteres magasság esetében a földfelszíni vágáslapon ez a hidrosztatikus nyomás 80 KPa (kilopascal). Mi tehát a teendő, hogy ettől a magasságfüggvényű, változó nagyságú hidrosztatikus nyomásból fakadó problémától megszabaduljon a növény? A nehézséget az okozza, hogy változó nagyságú nyomó-, illetve szívóerőre van szükség, és ennek legnagyobb szélső értékével még kis magasság esetében sem tud megbirkózni a fa.
Nézzük meg, hogy az épületgépészek hogyan juttatják fel az akár háromszáz méternél is magasabb felhőkarcoló felső emeletére a vizet. Elvileg két megoldás létezik. Egyenszilárdságú csövet hozunk létre egy nagyon erős vízpumpával és a szerelvényeket nyomáscsökkentőkkel védjük. Ilyen konstrukció tudomásunk szerint nem készült.
Az alkalmazott megoldás a vízvezetékrendszer leszakaszolása puffertárolók segítségével. A növény vízcsöve is ezen az elven működik. A leszakaszolás darabszámának arányában csökken a hidrosztatikus nyomás, de elérhető az is, hogy a szükséges oldalirányú erő oly mértékű legyen, ami lehetőséget teremt ahhoz, hogy általa a leszakaszolt csőben felfelé nyomás történhessen. A másik érdekes és fontos kérdés, hogy vajon ebben az esetben milyen pulzálási mozgásforma alakul ki? Vizsgáljuk meg, hol találunk leszakaszolásokat és ott milyen a mozgásforma. Az ember vékonybelében és érhálózatában ilyen a mozgás, de a gyűrűsférgek is hasonlóan mozognak, sőt a pocok is így mozog, mikor állkapcsát kiakasztva lenyeli a kígyó és továbbmozgatja bélrendszerében. Talán már sejtjük.
Perisztaltikus mozgásformával állunk szemben.
A fák vízcsövének esetében ez egyedi, sok tekintetben eltér a többi perisztaltikus mozgástól, de sok a hasonlóság is. A vékonybél perisztaltikus mozgásától annyiban tér el, hogy ott van egy szegmentációs (leszakaszoló) mozgás is, mely az idő függvényében előrehalad, és a megemésztett anyag forgatásában is szerepet játszik. A fák vízcsövénél szegmentációs mozgásra nincs szükség, mert azok szelepek által eleve leszakaszoltak. Vizsgáljuk meg ezeknek a leszakaszolt szegmentumoknak a mozgását, tárjuk fel a szelepek zárásának, nyitásának a mechanizmusát.
A szelepek zárásának, nyitásának a mechanizmusa
Az 1.A ábrán látható, ahogy a 2. számú szegmentumból vizet nyom a növény a felette lévő 1. számú szegmentumba. Fny=nyomóerő, Fsz=szívóerő, Fe=ellenerő (több erőből áll, ezért nem ábrázoltam)
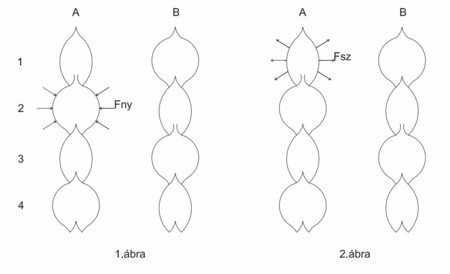
1. ábra: A szelepek zárása-nyitása 2. ábra: A szelepek zárása-nyitása
a nyomási szakaszban a szívási szakaszban
A szelepnyitás feltétele: Fny > Fe. A nyomási folyamat közben az ellenerőnek meg kell nőnie ahhoz, hogy a szelep bezáródjon. Abban a pillanatban, amikor Fny < Fe, záródhat a szelep. Ezután megkezdődhet a nyomás a 3. szegmentumból a felette lévő, előző nyomással lecsökkentett vízmennyiségű 2. számú szegmentumba (az eredményt az 1B ábra szemlélteti). A szelep nyitás-zárás előfeltétele egyezik az előbbivel. Ez a folyamat folyton-folyvást ismétlődik és megkapjuk az 1. ábrán látható perisztaltikus mozgásformát.
Az éjszaka lezajló gyökér felőli szívás mechanizmusát a 2. ábra szemlélteti.
Itt a szívás az 1A szegmentumból indul, ami vizet szív az alatta levő 2. számú szegmentumból. A szívás addig tart, amíg Fsz > Fe. Abban a pillanatban, amikor Fe > Fsz, a szelep bezáródik. A szívás további folyamata hasonlatos a nyomási folyamatban ismertetettekhez. Itt is kialakul a perisztaltikus mozgásforma, melynek eredményét a 2B ábra mutatja.
A nyomási folyamatban nappal valamennyi szegmentum egyre kisebb lesz, ami feltételezi azt, hogy lent a pulzálás kezdetén egy zárószelep helyezkedik el. Éjszaka a gyökér felőli szívásnál a lenti szelep kinyílik és az egész rendszer egyre vastagabb lesz, hiszen vizet szív. A pulzáló rendszer működése közben nekifeszül a belső (farész) és a külső (háncsrész) hengerpalásthoz, mert ebben az esetben a hidrosztatikus nyomásból fakadó erőt a két hengerpalást veszi fel, nem pedig az erre szilárdságtanilag alkalmatlan vékonyfalú pulzáló rendszer.
Mindezekből az is következik, hogy a külső hengerpalást, tehát maga a fa is változtathatja a méretét. Nappal kisebb, éjszaka az átmérője nagyobb lesz, tehát a háncsrész rugalmasnak tekinthető. (A fa átmérőváltozására vonatkozóan több irodalmi adat is van.)
Ezzel a perisztaltikus pulzáló mozgásformával már lehetősége van a csőben áramló víznek a legmagasabb fák csúcsába is feljutni.
A fák vízszállításának a fent leírt új elméletét először Török ismertette az Erdészeti Lapok 2016. évi januári és júniusi számában, a „Fák vízszállításáról újszerűen” címmel. A Forest Press elektronikus hírportálon 2015. július 28-án „A pulzáló rendszer – a fák vízszállításáról újszerűen”, majd 2016. február 29-én „A pulzáló rendszer féregmozgásai a fák vízszállításáról újszerűen” címmel írt cikket.
Bizonyítékok a perisztaltikus pulzálásra
• Béres Csilla izotópos kísérlete bebizonyította, hogy a fákban az áramlás nem egyenletes, hanem rövid ideig kiugró csúcsok jelennek meg, majd csökken a sebesség. Ez a mozgásforma éjszaka is tapasztalható volt, a csúcsok éjszaka is jelentkeztek. A kísérlet nem csak azt bizonyította, hogy az áramlás nem egyenletes, hanem egyúttal cáfolta a régi elméleteket is, mely szerint az áramlás mozgatóereje a víz párolgása. Éjszaka a sztómák zárva vannak, így a párolgás húzóerejével ez a jelenség nem magyarázható. (Béres Csilla cikkét az Élet és tudomány 1999/50. és 1999/51. számában ismertette „A fák vízforgalma” címmel.)
• A későbbiekben a kísérletet hordozható CT (komputertomográf) és MRT (magmágneses rezonancia tomográf) alkalmazásával is megismételték. Az elért eredményekről „A fafajok vízforgalmának összehasonlítása” címmel a VII. Természet-, Műszaki és Gazdaságtudományok Alkalmazása Nemzetközi Konferencián (Szombathely, 2008. május 17.) számoltak be. A kutatócsoport tagjai: Béres Csilla, Németh László, Zentai Zoltán, a NYME TTMFK Kémiai- és Környezettudományi Tanszék kutatói voltak.
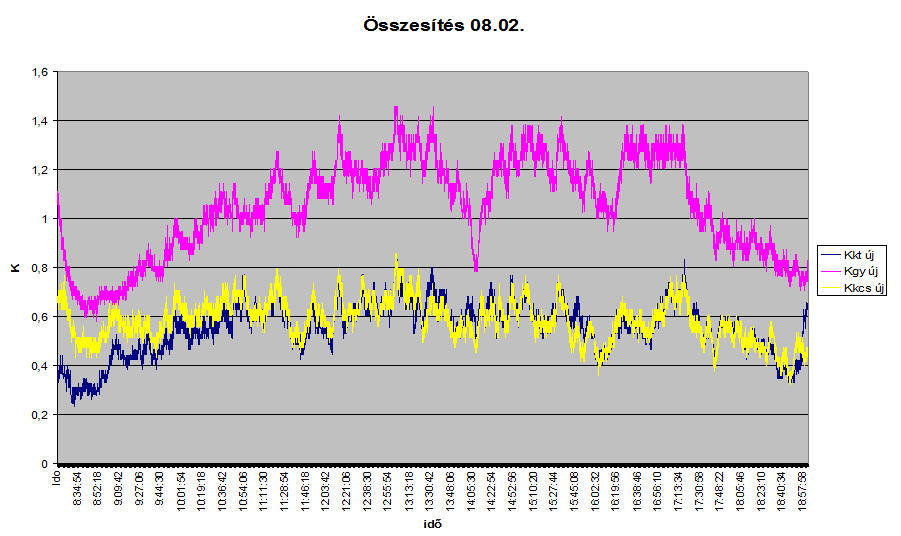
3. ábra: Csertölgy, kocsánytalan tölgy és gyertyán törzsében mért vízáramlás öntözés utáni napon.
A 3. ábrán a csertölgy, a kocsánytalan tölgy és a gyertyán törzsében mért időfüggvényű sebességmintázatok látszanak, egy locsolás utáni napon. A függőleges tengelyen K=0,206 U a 0,814 ezrediken, ahol U az áramlási sebesség (m/perc). Ugyanaz látszik, amit az izotópos kísérletnél is leírt. Az áramlás nem egyenletes, hirtelen gyorsabb és lassabb áramlások követik egymást. (Az időfüggvényű fűrész fogszerű sebességmintázat jól kirajzolódik a görbén.)
Vajon miért? A féregmozgásos pulzációs elmélet szerint a víz az egyik szegmentumból áramlik a felette lévőben. Kialakulhat-e itt egyenletes mozgás? Véleményünk szerint nem. Amikor a szegmentum oldalirányú ereje felnyitja a felette levő szelepet, akkor a víz meglódul, majd lelassul. Tulajdonképpen arról van szó, hogy a felfelé haladó víz kinetikus energiájának egy része elraktározódott a rugalmas szelepben, melyre hidrosztatikus és rugalmas rendszerek nyomása is hat, majd a szelep felnyílásakor ennek a kinetikus energiának egy része felszabadul, egy hirtelen gyors majd lassuló vízmozgásformát előidézve. A műszer által készített időfüggvényű sebességmintázat tehát visszatükrözi a szegmentumról szegmentumra áramló víz mozgásformáját.
• A kontinuitás törvénye önmagában is igazolná az elmélet helyességét, hisz a víz mozgása egy fogazott szinuszgörbéhez hasonlatos időfüggvényű sebesség mintázatot eredményezett. Nappal az átlagsebesség (v1) nagyobb, éjszaka pedig (v2) kisebb (a görbe éjszakai része hiányzik, alakjára következtetni lehet). Mivel a kontinuitás törvénye szerint v1*k1=v2*k2, ezért a mozgás nappal csak kisebb (k1), éjszaka pedig nagyobb (k2) keresztmetszeten következhet be. Tehát a cső pulzál.
• Gyakran lehet olvasni növényélettani könyvekben, hogy a gyökérnyomásnak van egy napi ciklusa és ez reggel a legmagasabb érték, mielőtt a gázcsere nyílások kinyílnak, és a párologtatás megkezdődik. Vajon miért?
Az éjszakai gyökér felőli szívás esetén a cső egész éjjel duzzad, nő a fa átmérője. A legnagyobb duzzadtságot és nyomást sztómanyitáskor éri el, amikor a cső elkezd felfelé nyomni. Gyökérnyomásról itt azért nem beszélhetünk, mert egy nyomóerő folyamatos mozgást nem tud előidézni, azt csak az erő által működtetett mechanizmussal lehet elvégezni.
• A fa életének egyik legrejtélyesebb titka: hogyan indul be tavasszal az élet, hogyan lesz a rügyben szunnyadó hajtáskezdeményből leveles hajtás? Ne felejtsük el, hogy ekkor még sem párolgás, sem fotoszintézis sincs.
Véleményünk szerint ősszel a színesedő levelek úgy esnek le a fáról, hogy a nyomás benne marad a rugalmas szállítórendszerben. A fáknak nem is áll módjukban másként lezárni a csövek végét, csak nyomás alatt, hisz a pulzáló rendszer, ha csökkenő mértékben is, de felfelé nyomva működik. Tél végén, amikor ez a nyomás a vízcsövekből kiszabadul, azt tapasztaljuk, akár a legmagasabb ágak lemetszése esetében is, hogy a nyomás alatt lévő cukros többletvegyület kifolyik a fából. Ez a nyomás és többletvegyület az, ami az élet újraindulásának nélkülözhetetlen előfeltétele. A vízcső mellett cukros vegyületek találhatók, melyek rügyfakadás előtt egyesülnek a csőből kiszabaduló oldattal.
• Szakirodalmi adatok bizonyítják, hogy a fák változtatják átmérőjüket. Éjszaka a szívási fázisban nagyobb, nappal a nyomási szakaszban pedig kisebb az átmérő értéke.
• Egy magyar-finn-osztrák kutatócsoport infravörös lézerszkennerrel vizsgálta, mi történik a fák lombjával, ha leszáll az éj. A tudósok arra jutottak, hogy a fák is alszanak. A vizsgált fákról lézerszkenneléssel nagy pontosságú és részletességű modellt készítettek és ezt az éjszaka folyamán óránként ismételték. Beigazolódott, hogy a fák ágai és levelei éjszaka akár tíz centiméterrel is lejjebb ereszkednek. Zlinszky András az MTA biológusa szerint napkelte előtt érték el a legalacsonyabb pozíciójukat, majd hajnalban néhány óra alatt visszatértek korábbi helyzetükbe. Egyelőre nem tudják, hogy a nap ébresztette-e fel a fákat, vagy a saját belső ritmusuk, a naptól függetlenül – olvasható a kutatásvezető Eetu Puttonen, a Finn Geodéziai Kutatóintézet munkatársának beszámolója a kutatási közleményben (forrás: MTI/Szerk. on 2016-06-03 at 14:00).
Azt gondoljuk, hogy a fák alvómozgásánál szemléletesebben semmi sem bizonyíthatja az elmélet helyességét. Éjszaka a gyökér felőli szívás esetén a kihajló ágak a szállítórendszerbe áramló víztől egyre nehezebbek lesznek, így folyamatosan lejjebb ereszkednek. Érdekes kérdés: hajnalban miért nyerik vissza néhány óra alatt eredeti helyzetüket, hiszen nappal a párolgás hatására a csövekben tárolt vízkészlet egész nap, nem csak néhány óráig csökken. Ez a jelenség a rugalmas rendszerekben a víz nyomásának hatására bekövetkezett alak- és helyzetváltozással magyarázható. Napkeltekor a cső elkezd felfelé nyomni. Ez a nyomás képes az eredeti helyzetet rövid időn belül visszaállítani. Egy lezárt végű gumicső, amelyben kis nyomás van, függőlegesen a kézbe tartva ívesen meghajlik. Nyomás alá helyezve gyorsan felemelkedik, a nyomás mértékétől függően közel a függőleges helyzetig is, ha nem túl hosszú a gumicső.
Epilógus
Befejezésül fontosnak tartjuk, hogy néhány gondolat erejéig érintsük a bionikát, ezt a modern tudományt, hisz írásunkban oda-vissza élő és élettelen rendszereket hasonlítottunk össze. A bionika olyan új, több szaktudományt átfogó tudományág, amelynek célja az élő természetben kifejlődött megoldások átültetése a műszaki gyakorlatba, abból a megfontolásból is kiindulva, hogy a természetben fennálló természetes kiválasztódás az optimális megoldásokat jelenti egy-egy problémára.
A bionika fő alapelve: hasonló rendszerek hasonlóképpen működnek. Oka vélhetően a mechanizmusuk működési hasonlóságában keresendő. A mechanizmus olyan önmagában záródó „szimmetrikus” mozgásforma, melyet az ellentétesen működő hatások tartanak szimmetrikus állapotban.
A fák organikusnak tekinthető vízcsövénél több ellentétes hatás is fellelhető: szívás-nyomás, keresztmetszet-növekedés – keresztmetszet-csökkenés, felmelegedés-lehűlés, szelepnyitás-szelepzárás. Ha a felsorolt ellentétpárokból egy is kimarad, vagy az ellentétpárok ellentéteiből egy is hiányzik, akkor az organikus rendszer mechanizmusa működésképtelenné válik. A vizet szállító vízszivattyúknál ilyen sok ellentétpárról nem beszélhetünk. A vízszivattyút nem készítette fel az ember arra, amire a növény vízszállító rendszerének fel kellett készülnie. Alkalmazkodni a változó klímához, a változó diszponibilis vízkészletekhez. Ezért az organikus rendszerek mechanizmusai sokkal kifinomultabbak. Nem szeretnénk abba a tévedésbe esni, ami azt sugallná, hogy a vízcső működése csupán fizikai és mechanikai törvényszerűségeken alapszik. A vízcsőnek a hideghatásra történő összehúzódása mögött bonyolult élettani, biokémiai és biofizikai folyamatok húzódhatnak meg. Úgyszintén élettani jelenségek irányíthatják a víztározó mozgásokat, amikor is az élő sejtekből felépült cső az élettelen, már elhalt vízcsövekben vizet tárol, és onnan vizet vesz fel. Ezeknek a jelenségeknek sem ismerjük élettani hátterét, ugyanúgy, ahogy az általuk irányított biokémiai folyamatok is ismeretlenek számunkra. Elmondhatjuk tehát, hogy a vízszállítás mozgatórugója a pulzáló rendszer mozgása, melynek hátterében élettani folyamatok állhatnak, de mozgásukat a fizika törvényeinek megfelelően végzik.
Végezetül csupán egy gondolat. Hogyan is szállítják a fák a vizet? Eltolt időfázisú szívással és nyomással, perisztaltikusan pulzálva. És mi emberek, hogy szállítunk? Ugyanígy. Eltolt időfázisú szívással és nyomással, perisztaltikusan pulzálva. Talán nem véletlenül.
Összefoglalás
A fákban történő vízszállításnak az az elmélete, mely szerint a víz szállításának legfontosabb hajtóereje a víz párolgása következtében a párologtató elemeken megjelent meniszkuszok (a kapilláris folyadékoszlop görbült felszínei) által létrehozott szívás, melyet alulról a gyökérnyomás is megtámogat, hibás.
Hibás, mert:
- A rendszerben nincs elegendő erő: az ismert kapillárishatással az emelés egy méter körüli érték a természetben. A gyökérnyomás vízszállításra alkalmatlan, mert az erő csak az általa működtetett mechanizmussal képes folyamatos munkavégzésre, például vízszállításra.
- Hibás a mechanizmusa. Vizet szállítani csak úgy lehet, hogy a felszívott vizet kinyomja a rendszer (szivattyúknál ez történhet egy időben vagy késleltetve). Vizet szállítani úgy nem lehet, hogy alul nyomok, felül szívok, és úgy sem, hogy csak szívok, vagy csak nyomok.
- Egy középmagas bükk húsz méteres magasságban lévő levelébe két atmoszféránál egy kicsivel nagyobb szívóhatás keletkezik. Könnyen belátható, hogy egy ilyen nagy szívóerő a levél és ezzel együtt a fa azonnali pusztulásához vezetne. Egy rendszer akkor működik a célnak megfelelően, ha működőképes a mechanizmusa, létezik ehhez illeszkedő megfelelő nagyságú és állandóan ható erő úgy, hogy szerkezeti elemei ezt képesek elviselni. Ezeket a feltételeket a régi rendszer nem tudja teljesíteni.
Ha nem így, akkor hogy szállítja a fa vizet?
Párolgáskor a lombkorona irányába nem szívás, hanem nyomás történik, a párolgás miatti hőelvonás következtében előállt vízcső keresztmetszet-csökkenése miatt. Éjszaka a párolgás szünetelésekor beindul egy hőkiegyenlítődési folyamat, ami visszaállítja a cső eredeti keresztmetszetét, ami szívást generál és felszívja a vizet a talajból. Ekkor a rugalmas cső teljes keresztmetszete pulzál, ami lehetővé teszi az állandó erőhatás miatti szállítást a csőben. Mivel a csőben nyomásgrádiens alakul ki, ami változó nagyságú, nagyobb famagasság esetén meg tetemes erőt igényelne, a rendszer leszakaszolt. Ez pedig egy perisztaltikus mozgásformát eredményez.
A cső tehát eltérő időfázisban szívó-nyomó perisztaltikus mozgást valósít meg. Ennek bizonyítását több jelenség is igazolja.
A kontinuitás törvénye önmagában is bizonyítja az elmélet helyességét, hisz a víz mozgása egy fogazott szinuszgörbéhez hasonlatos időfüggvényű sebességmintázatot eredményezett. Nappal a (v1) átlagsebesség nagyobb, éjszaka pedig (v2) kisebb. A kontinuitás törvénye értelmében (v1*k1=v2*k2), a mozgás nappal kisebb (k1), éjszaka pedig nagyobb (k2) keresztmetszeten következhet be. Tehát a cső perisztaltikusan pulzál. A víz szállítása azonban nem csupán fizikai törvényeken alapszik. A vízcsőnek a hideg hatására történő összehúzódása mögött bonyolult élettani, biokémiai és biofizikai folyamatok húzódhatnak meg. Török András
1